Introduction
SDS polyacrylamide gel electrophoresis (SDS PAGE) will be used to assess the purification process and to determine the apparent molecular weights of the three apoproteins.SDS-Polyacrylamide gel electrophoresis
Electrophoresis is the process in which charged particles migrate through a solid or liquid matrix in response to application of an electric field. Rate of particle movement is proportional to the charge:mass ratio of the particle and to its frictional resistance. Larger particles move more slowly, and highly charged particles move more quickly. In protein electrophoresis, these factors tend to balance out. Size and charge of a protein determine its electrophoretic mobility. If proteins are separated through a gel matrix with varying pore size, migration depends on the size and shape of the protein. Smaller proteins are retained less, and thus move faster. On the other hand, the net charge of a protein depends on the pH. In native gel electrophoresis, both charge and size determine the migration pattern; in this technique excellent separation can be achieved, but unambiguous information about the protein size cannot be obtained.
However, proteins which have a similar net charge separate nicely according to their size, provided they are of similar shape. For example, globular proteins are retained less than rod-like proteins of the same molecular weight. Experimentally, we can eliminate the influence of charge on protein migration by providing all proteins with extreme negative charges. Sodium dodecyl sulphate (SDS), is an anionic detergent that binds to proteins. Its action is to denature the protein by solubilizing it and effectively "coating" it with a negative charge. The effect before electrophoresis is to block protein interactions with other proteins, polymers such as nucleic acids and lipid, to dissociate multimeric proteins; and to alter folding in protein monomers. SDS not only provides proteins with a strong negative charge, but it also denatures the protein, thus giving each protein a roughly globular shape; differences in protein shape that would affect electrophoretic mobility are eliminated.

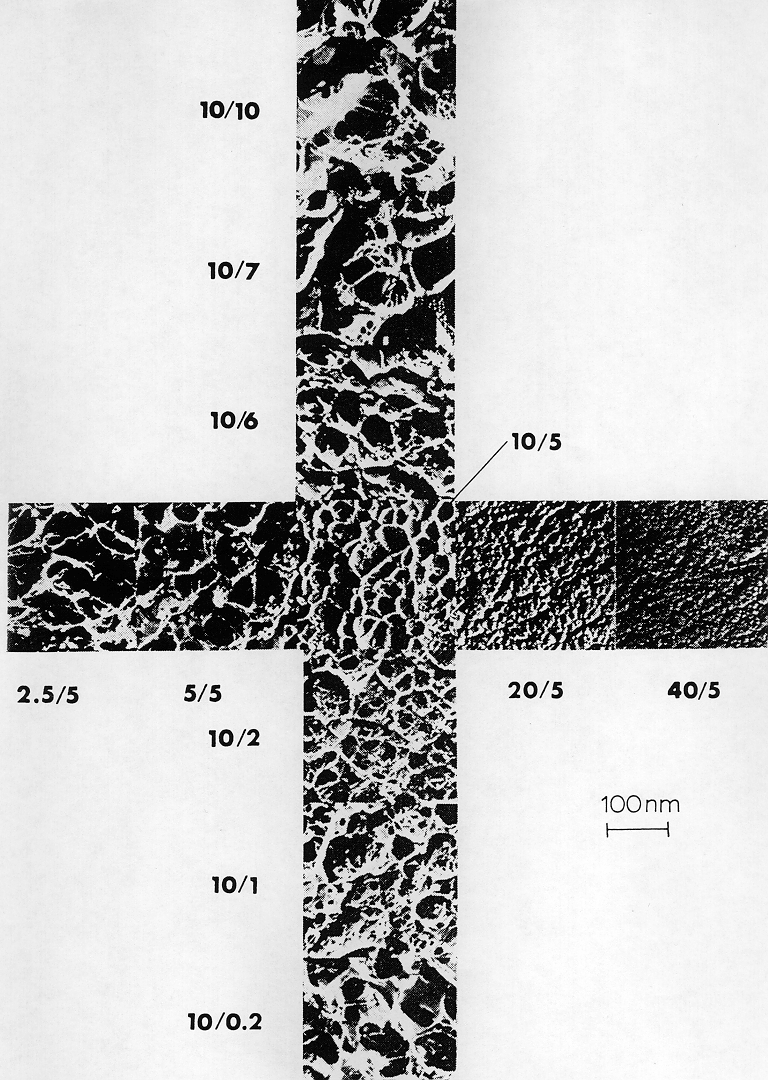
The gel matrix most commonly used for proteins separation is polyacrylamide. Polyacrylamide gels are formed when monomeric acrylamide is polymerized by the action of a radical forming agent, ammonium peroxidisulfate (ammonium persulfate) and N,N,N',N'-Tetramethylenediamine (TEMED). The gel forms optimally in the absence of free oxygen, since oxygen is a stable di-radical which can terminate the radical induced polymerization reaction. Since acrylamide polymerizes to long linear products, a cross-linker is required to form a three-dimensional gel. Bisacrylamide serves this function. Pore size within the gel is determined by both, the total acrylamide concentration (% T; = g acrylamide + g bisacrylamide per 100 ml) and the relative concentration of the cross-linker bisacrylamide (% C; = g bisacrylamide per 100 g (acrylamide + bisacrylamide). While pore size decreases with increasing T, small and large C-values yield large pores; the smallest pores are formed in the presence of approx. 5 % C, as seen in this electron micrograph.
{kind=link}

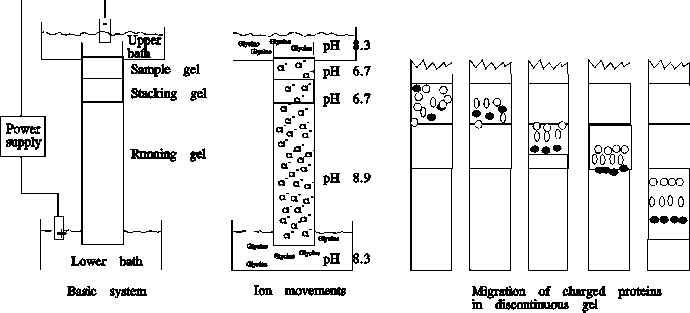
Gels can be cast as columns or slabs. For analytical purposes, slabs are much more widely used, since they allow the separation and comparison of multiple samples. In either case, a "stacking" gel is placed on the top of the separating ("running") gel to sharpen the bands before they enter the gel. The electrophoresis buffer and the buffer in the separating gel have a high pH (8.9) and contain glycine. In contrast, the stacking gel buffer has a low pH (6.8) and contains Cl-. At the low pH of the stacking gel, the Cl- in the stacking gel are negatively charged and hence move towards the anode (+), but the glycine entering from the gel buffer has only a very small negative charge (pI of glycine ~ 6). Thus, Cl- moves faster than glycinate, and within the stacking gel a zone of low anion concentration (= low conductivity) forms. This leads to a higher electric field, which accelerates the proteins so that they enter the separating gel as a narrow band at the boundary between the leading Cl- and the trailing glycinate ions. When the protein complexes reach the running gel (pH 9.8), the glycine becomes completely dissociated, and migrates at the same speed as Cl-. You can look at a schematic illustration of this mechanism.
{kind=link}
Source: http://www.bioon.com.cn/protocol/showarticle.asp?newsid=5812
0 comments:
Post a Comment